|
LA GENETIQUE
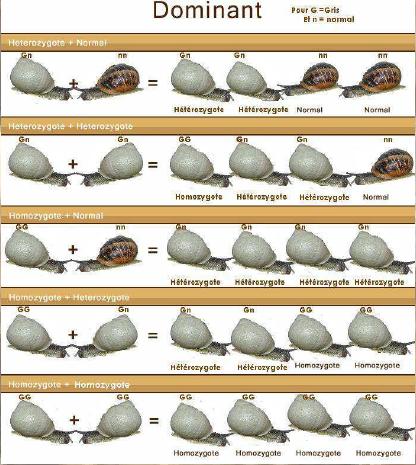
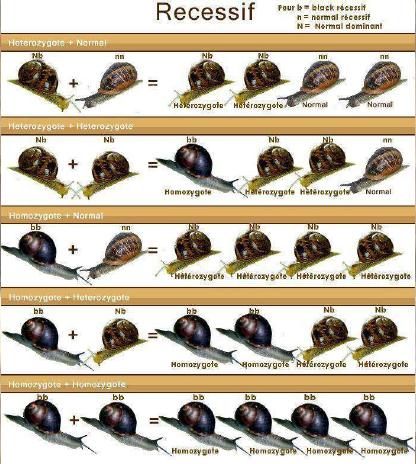
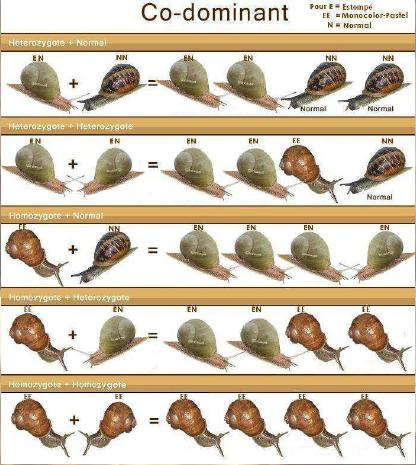
Dernière mise à jour: 19/06/2008 Rappel: HELIX ASPERSA: n = 27 Chromosomes, 2n = 54 Chromosomes Le génotype
d'un individu est l'ensemble des allèles (exemplaire,
version d'un gène) d'un individu. - si ces 2 allèles sont identiques, l'individu est homozygote (pour ce gène) : il est A//A ou B//B - si ces 2 allèles sont différents, l'individu est hétérozygote (pour ce gène) : il est A//B Un gène peut avoir plusieurs allèles, c'est le poly allélisme. Si il possède de très nombreux allèles, il est polymorphe. Si l'organisme est homozygote, son phénotype est celui déterminé par l'unique version du gène qu'il possède. Si l'organisme est hétérozygote, son phénotype peut soit être déterminé par un seul des deux allèles (on dit alors que cet allèle est dominant, l'autre étant qualifié de récessif), soit correspondre à l'expression des deux allèles à la fois (on dit qu'il y a codominance entre ces allèles). Il n'y a pas de chromosomes sexuels chez ces animaux. En tout cas, pas de chromosomes qui différencieraient des cellules mâles ou femelles... Par contre il y aurait des gènes qui s'exprimeraient de manière différentes suivant les cellules où le moment de la vie de l'animal... Ce principe n'existe pas que pour la différenciation des gonades. Bref, il s'agit plutôt d'un différentiel d'expression génique que d'une structure particulière dans les chromosomes. Variation acquise et variation génétique: Les facteurs susceptibles d'influencer la structure génétique de la population sont les forces évolutives telles que la migration, sélection, mutation et dérive génétique ainsi que les contraintes inhérentes au régime de reproduction c'est-à-dire absence de croisement au hasard, recouvrement des générations. Les caractères d’un individu sont en partie la conséquence des conditions extérieures qu’il subit à l’instant considéré, ou qu’il a subies dans le passé au cours de son développement. L’existence de ces caractères que l’on qualifie d’acquis est très manifeste chez les organismes supérieurs. Des traumatismes subis au moment de la naissance, peuvent perturber gravement et définitivement le développement de l’adulte. Les caractères acquis ne sont pas transmis à la descendance. Les caractères acquis ne correspondent qu’à des variations dans l’expression des potentialités de l’organisme et seules celles-ci sont héréditaires. Nous avons plusieurs moyens d'effectuer une sélection d'élevage ; chacun pourra choisir ce qu'il pense le mieux adapté pour la programmation des accouplements : - au hasard - par homogamie (réunir les meilleurs) - par exogamie (les meilleurs avec les moins bons pour améliorer la descendance) - par endogamie : la consanguinité (large ou étroite) Sur les exemples de croisement suivant (photomontages), le propos n’est pas d’aboutir à une coloration de coquille quelconque, il s’agit plutôt d’illustrer la transmissibilité de caractères particulièrement intéressants en élevage, par exemple : prolificité des pontes, poids adulte, etc.….
La consanguinité surtout étant à l'ordre du jour, voici quelques explications : C’est une méthode d'élevage ; elle n'est pas responsable des bons et des mauvais résultats. Ceux-ci dépendent avant tout de la façon dont on la pratique. Elle consiste à accoupler des sujets de grande qualité, ayant un ou plusieurs ascendants communs ; on tente de retrouver leur qualité dans la descendance par le phénotype des géniteurs de départ. L'espèce étant dotée d'une grande variabilité, il n'y a pas de sélection réfléchie sans consanguinité, sinon le programme d'élevage s'apparente au tirage du loto. De plus, il faut comprendre que la sélection consanguine va jusqu'à l'accouplement en 4e ou 5egénération. Au delà, l'effet de dilution des gènes est trop important. Il est impératif de s'assurer de la qualité des géniteurs de départ mais aussi des intermédiaires, la consanguinité devant être pratiquée sur plusieurs générations pour être utile. Ce type de sélection n'apportera jamais plus que les caractères apportés par les géniteurs de départ. On peut et on doit éliminer dans la descendance certains des défauts de famille dès qu'ils apparaissent, mais pour les qualités, il est impossible de dépasser un certain seuil, une fois qu'il est atteint. Toute interruption de la consanguinité interrompt la sélection et mène à l'hétérozygotie. A l'inverse, une sélection trop étroite constitue une erreur dangereuse, les résultats ne pouvant être que lents et progressifs. L'éradication d'un défaut ou même d'une tare ne peut se faire sur 2 ou 3 générations. L'hérédité quantitative est trop complexe pour permettre d'obtenir une expression constante des caractères recherchés ou non souhaités. Endogamie : Croisement d’individus avec un grade parental proche (issus d‘une même ponte, parents collatéraux, etc.) Consanguinité. La consanguinité de ces issus entre eux est en moyenne la même que celle vis-à-vis de leurs parents. Cela implique qu'un escargot issu d'un accouplement d’une même fratrie a le même degré de consanguinité qu'un escargot issu d'un accouplement entre parent et enfant. La multiplication de ce type d'accouplement conduirait rapidement, sans l'apparition aléatoire de mutation parfois significative, à une consanguinité de 100%, c’est-à-dire à un clonage avec un unique allèle pour chaque caractère non sexuel : au départ, chaque gène dispose d'au maximum 4 allèles différents (un pour chacun des deux chromosomes de chacun des deux parents), et à chaque accouplement chaque copie à une chance sur deux de disparaître. Mais, inversement, il suffit d'un seul accouplement exogame pour faire immédiatement tomber la consanguinité à 0% conduit à l’homozygotie. Exogamie: Croisement d’individus d’un grade de parenté éloigné. Land Breeding. Conduit à l’hétérozygotie. Et pour savoir si un escargot de phénotype dominant (= apparence), par exemple monocolore, est porteur de dilution (donc hétérozygote), il faut le marier avec un homozygote récessif. Ce mariage s'appelle un back cross ou croisement en retour. C'est le seul mariage permettant de préciser ce que porte un hétérozygote éventuel. En effet, un homozygote ne produit qu'une sorte de gamètes (pour un caractère, cela va de soi), ce qui mettra en évidence les différents gamètes produits par un hétérozygote. Vigueur Hybride : Elle se produit après appariement de deux lignées, où la F1 est plus productive que celle de ses géniteurs. La vigueur hybride, se traduit par un gain de performances qui résulte du mélange de différents gènes de différentes races, une « stimulation physiologique » due à l'état hétérozygote. Sélection par générations séparées : On emploie chaque génération et on la soumet à une sélection et un appariement. Ce type de méthode engendre un intervalle génératif plus ample. Sélection par générations imbriquées : les reproducteurs de toutes les générations sont ajoutés au processus. L'appariement et une sélection phénotypique sont continuels. (Il y a des animaux de tous les âges). Types d'appariement : Ils permettent de composer des caractéristiques sélectionnées au préalable et être imprimées aux futurs reproducteurs. Les méthodes de sélection nous indiquent comment sera le but du processus, mais le système d'appariement est le moyen pour arriver au but cherchée : • Au hasard • Par relations familiales • Par relations phénotypiques Toutes améliorations génétiques impliquent deux processus "sélection"-"Appariement ". Par le type d'individus (hermaphrodites incomplets) dans l’héliciculture le meilleur système d'appariement est basé sur l'hétérosis (L'hétérosis désigne l'augmentation des capacités et ou de la vigueur d'un hybride par rapport aux races, lignées... dont il est originaire) et les relations consanguines. Ce processus est massif et il s'améliore sur l'amélioré. Les croisements les plus d'employés sont ceux qui ont pour but obtenir une croissance plus élevée, puisqu'il est connu que quand deux individus de lignes distinctes se croisent une plus grande vigueur se produit en ce qui concerne le ou les caractères sélectionnés " la Vigueur Hybride ". Le système de sélection à utiliser est dénommé "Sélection réciproque récurrente modifiée". C'est une méthode de sélection et de reproduction combinée. Elle est basée sur l'existence, le développement et combinaison de deux souches, lesquelles seront choisies par le progrès génétique de la progéniture, à son tour de cette progéniture on choisit les meilleurs individus quant aux caractéristiques phénotypiques, puisque les caractéristiques génotypiques des mollusques terrestres (escargots Helix aspersa) sont encore inconnues.
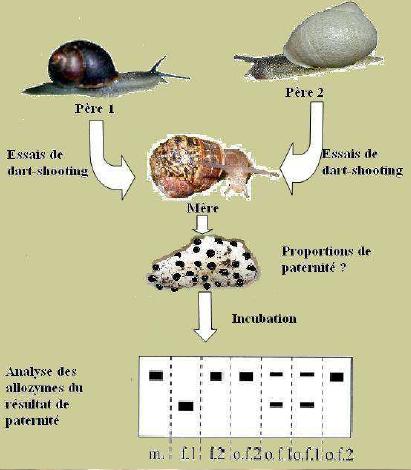
Conception expérimentale pour évaluer la relation entre l'efficacité du « dart-shooting » et le succès reproducteur paternel pour H. aspersa. le m, la mère; f1, père 1; f2, père 2; of1, résultat de père 1; of2, résultat de père 2. Les caractéristiques phénotypiques à sélectionner seront : chez l'ascendant, comme première mesure la quantité de progéniture qu'il développe (ponte, mortalité embryonnaire et post natale) et les autres se baseront sur les caractéristiques prises par la descendance proprement dite, dans laquelle seront vus des facteurs tels qu'AMPV (une Augmentation Mensuelle de Poids vivant), pigmentation de la chair et de la coquille, vivacité, dominance ou non à l'intérieur de la population, etc. Tous ces facteurs influeront sur la pression de sélection qui est prise. Avec ces valeurs pour indice on trouve une moyenne statistique et on sélectionne les individus qui sont sur ou très proches de cette moyenne choisie. Ces individus sélectionnés (F1) feront partie des sujets potentiels à sélectionner. Une fois arrivé à un degré proche de parenté on introduit une nouvelle souche d’un autre lieu (sélectionnés par les mêmes caractéristiques phénotypiques) et ils sont soumis à une reproduction en obtenant une plus grande progression productive donnée par la vigueur hybride et / ou l’hétérosis multiple. La génétique de la rotation de la coquille d'escargot: La fréquence à laquelle surviennent les escargots à coquilles dextres et sénestres dans la nature, va dans le sens d'un simple schéma dominant-récessif de l'hérédité. La forme sénestre est clairement le caractère ou allèle récessif, réprimés dans un génotype hétérozygote par le caractère dominant de type sauvage, selon Mendel. Certaines expériences de croisements, donnent des résultats plutôt inattendus, ce qui en aucun cas explique le nombre étonnamment faible de coquilles sénestres dans la nature, que l’on dénombre à une seule pour plusieurs milliers de dextres (1/20 000). Selon le modèle d’hérédité normale dominant-récessif, les escargots qui portent l'allèle récessif, pour une coquille sénestre homozygote, donc dans la combinaison ss, s provenant des deux parents seraient les seuls sénestres. En accouplant deux d'entre eux, la génération de leurs enfants (la génération F1) devrait être exclusivement sénestre, puisqu’il n’y a pas d’allèle D (dextre) qui apparaît. Contrairement, leur progéniture sera plus que probablement, et presque exclusivement, dextre : 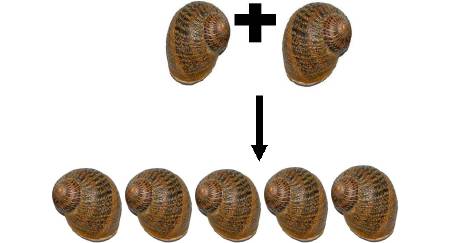 En se basant sur un modèle dominant-récessif de l'hérédité, c’est inexplicable !!! L'explication a été trouvée par le généticien américain Henry Alfred Sturtevant en 1923, en utilisant un escargot aquatique. Il existe un effet dominant-récessif, et aussi un effet matrocline (relatif à un individu ayant surtout hérité génétiquement de sa mère). Cela veut dire que, la rotation d’une coquille d'escargot n’est pas influencée par son génotype (la combinaison de ses allèles), mais par celui de sa mère. Ce fait étonnant est provoqué par les escargots appartenant au groupe d'animaux Spiralia. Leurs cellules se fécondent dans une spirale. Le sens de rotation de cette spirale, et donc finalement, l'enroulement de la coquille, sont déterminés par la direction de l'information génétique dans le cytoplasme de la cellule. Comme la cellule est une cellule oeuf, c'est l'information génétique de la mère qui prime, et non celle de sa progéniture. D’où cette cinquième possibilité. Voici donc le même schéma de croisement, cette fois avec aussi chaque génotype. (D: dominant, (dextre); s: récessif (sénestre). 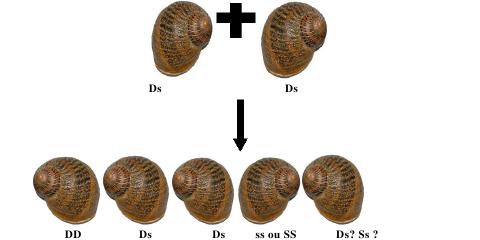 Le cinquième escargot est hors des théories de Mendel, la relation entre les génotypes DD: Ds: ss étant généralement 1:2:1. Un seul de ces escargots de cette progéniture, s’il survit jusqu'à maturité sera sénestre : C'est celui avec le génotype ss. En toute vraisemblance, le caractère "sénestre" disparaîtra à nouveau dans la prochaine génération, réprimés par les gènes dominants du père. Par ailleurs, seuls 5% des jeunes escargots survivent dans la nature, c’est une autre raison importante de la rareté des sénestres. Voici un schéma de croisement pour l'un des nombreux cas : 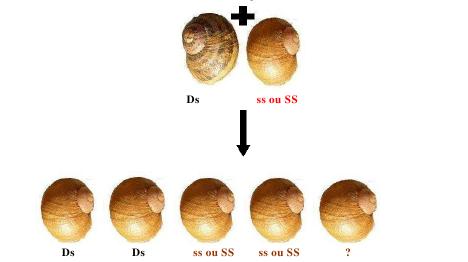 Quel
pourrait être le génotype des rejetons 3,4, ainsi que pour
le génotype possible de la cinquième possibilité
dans ce dernier schéma de croisement.(ne cherchez pas, lisez
plutôt la suite) Il se
trouve que les escargots peuvent uniquement se reproduire avec des
individus de même chiralité : en effet, les organes
génitaux sont orientés. Du coup, un escargot dextre ne
peut pas copuler avec un escargot sénestre : les individus
“DD” et “ss” ne peuvent se reproduire entre
eux, et forment donc deux espèces différentes…
Si dans notre population a chiralité droite, imaginons qu’un individu “Ds” apparaît. Lui peut se reproduire avec les autres de chiralité droite et transmettre l’allèle s, dans sa descendance, on trouvera des “Ds”. A un moment, il peut arriver également que certains de ses descendants copulent et génèrent des individus “ss”, qui seront alors sénestres. Seulement, la majorité de la population restera dextre, donc il leur sera difficile de se reproduire, même s’il peut y avoir d’autres individus sénestres… L‘orientation de la coquille n’est pas déterminée par le génotype de l’individu, mais par celui de sa mère ! Si une mère a génotype “ss”, les enfants seront sénestres, quel que soit leur génotype. Dans tous les autres cas, les enfants seront dextres. La direction de l'enroulement, commence avec la première divisions de l'œuf lorsque, le fuseau mitotique est incliné vers la droite ou vers la gauche. En résumé, un modèle émerge de ces données, selon lequel le fuseau agirait comme une sorte de boussole intracellulaire. Tout d’abord, comme l’aiguille de la boussole, le fuseau est asymétrique. L’asymétrie de la boussole est magnétique, celle du fuseau est chimique. Dans le cas de la boussole, l’asymétrie de l’aiguille lui permet de s’orienter dans un champ magnétique. Pour le fuseau, son asymétrie lui permet de s’orienter grâce à une molécule motrice dans un champ de câbles d’actine, si ces câbles sont eux-mêmes orientés. Le fuseau semble donc fonctionner comme une sorte d’organe sensoriel, qui est capable de percevoir l’orientation de la cellule et de transmettre cette information à l’appareil de division. La génétique de la couleur de la coquille de l'hélix aspersa: Le document est ici !!!! Les gènes du polymorphisme de la coquille de l'hélix aspersa: Le document est ici !!!! REFERENCES : SPECIATION AND GENE FLOW BETWEEN SNAILS OF OPPOSITE CHIRALITY http://biology.plosjournals.org/archive/1545- 7885/3/9/pdf/10.1371_journal.pbio.0030282-L.pdf LE FUSEAU MITOTIQUE EST-IL UN ORGANE SENSORIEL DE LA CELLULE ? Yves Barral, Institut de Biochimie, école polytechnique fédérale de Zurich, ETH-Zentrum Universitatstrasse 16, 8092 Zurich, Suisse http://www.erudit.org/revue/ms/2003/v19/n11/007273ar.html COMPARTIMENTATION ET DYNAMIQUE CELLULAIRES- UMR 144 CNRS / INSTITUT CURIE Equipe : Polarité cellulaire chez la drosophile Chef d’équipe : Dr. Yohanns Bellaiche, CNRS, DR2 http://www.curie.fr/upload/recherche/equipes/eq_281/umr144-cnrs-curie-yb-fr.pdf INHERITANCE OF DIRECTION OF COILING IN LIMNAEA A. H. STURTEVANT Columbia University http://www.esp.org/foundations/genetics/classical/holdings/s/ahs-23a.pdf SINISTRAL SNAILS AND GENTLEMEN SCIENTISTS J.B. Gurdon1,* 1Wellcome/Cancer Research UK Institute, Cambridge CB2 1QN, United Kingdom *Contact: jbg.1000@hermes.cam.ac.uk DOI: 10.1016/j.cell.2005.11.015 http://download.cell.com/pdfs/0092-8674/PIIS0092867405012237.pdf
|
|||||||||||